")
")
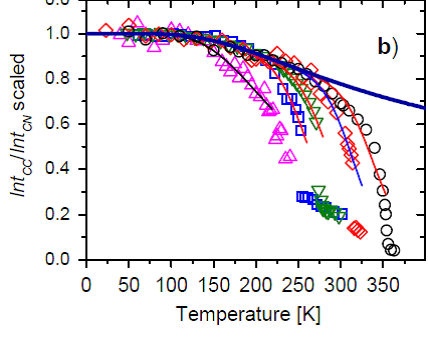
Научные и прикладные результаты лаборатории 0420171. Исследование спектров комбинационного рассеяния света (КРС) фосфолипидных мембран DMPC и DLPCРанее для пяти различных фосфолипидов было показано, что температурная зависимость интенсивности моды около 1130 см-1, измеренная методом комбинационного рассеяния света (КРС), хорошо описывает экспериментальные данные и позволяет определять энергию активации разупорядоченного состояния и ее степень вырождения. На Рис. 1.1 показана температурная зависимость отношения интенсивности линии КРС вблизи 1130 см-1 (IntCC) и интенсивность моды C-N(CH3)3 (IntCN, вблизи 720 см-1), служащей в качестве референтной линии. На этом рисунке данное отношение нормировано на свое значение в пределе низких температур, чтобы можно было сравнить поведение разных липидов.
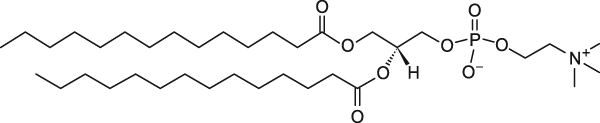
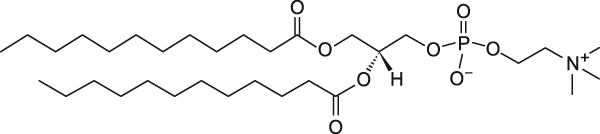
Рис. 1 Все фосфолипиды, исследованные ранее (Surovtsev N.V., Dmitriev A.A. // J. Phys. Chem. 2015), имели от 16 атомов углерода в каждом хвосте и больше. А что будет, если рассмотреть случай липидов с меньшим числом атомов углерода, но которые еще способны образовывать фосфолипидные бислои, служащие моделью мембранных структур в биологической клетке? В 2017 году мы рассмотрели случай насыщенного липида DMPC c 14 атомами углерода в хвосте и DLPC с 12 атомами углерода в хвосте.
DMPC
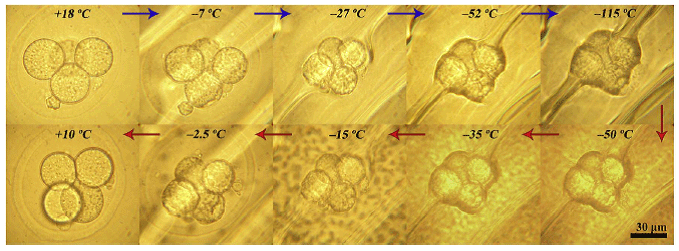
DLPC Рис. 2 Были приготовлены водные суспензии многослойных фосфолипидных везикул. Процедура синтеза включала в себя стадию «набухания» на температурах выше перехода гель-флюид и многократные (5-10 раз) процедуры закалки-отогрева, когда образец пересекал температуры перехода гель-флюид и плавления/замораживания льда. Ранее нами было показано, что такой протокол синтеза приводит к образованию многослойных везикул с размером в несколько микрон и числом слоев порядка сотни или сотен. Методом дифференциальной сканирующей калориметрии (ДСК) образцы были охарактеризованы и полученные температуры перехода гель-флюид (+24 С для DMPC и -2 С для DLPC) хорошо совпали с известными литературными данными. Также из эксперимента по ДСК были определены значения энтальпии для перехода гель-флюид, которые находятся в хорошем согласии с данными других авторов. Оказалось, что фосфолипиды с короткими хвостами также в целом следуют наблюдаемой корреляции между значением активационного барьера для разупорядоченного состояния хвостов фосфолипида и значением энтальпии перехода гель-флюид. 2. Исследование спектров КРС замораживаемых преимплантационных эмбрионов мышиИсследовалась проблема детектирования фазовых переходов липидов в замораживаемых эмбрионах мыши методом КРС. Криоконсервация преимплантационных эмбрионов и ооцитов ‑ один из наиболее технологичных и перспективных способов решения проблемы сохранения генетического разнообразия видов млекопитающих. В настоящее время криоконсервация эмбрионов и ооцитов активно используется в репродуктивной медицине, а также для криоархивирования лабораторных и сельскохозяйственных животных. Особо следует отметить применение криоконсервации эмбрионов и ооцитов для сохранения редких и исчезающих видов дикой фауны. Не смотря на то, что для некоторых видов животных выработаны эффективно работающие протоколы криоконсервации, для подавляющего большинства видов млекопитающих таких стандартных протоколов замораживания все ещё нет. Повреждающие факторы протоколов криоконсервации связаны с фазовыми переходами, возникающими при глубоком охлаждении. В то время как применяемые криопротекторы позволяют справиться с проблемами, вызываемыми кристаллизацией воды, фазовые переходы липидов, происходящие в липидных гранулах и мембранных структурах клетки, по-видимому, и ответственны за трудности, возникающие при криоконсервации ооцитов и эмбрионов «проблемных» видов животных (например, представителей отряда хищных). Повреждающая роль липидов отражается в известной корреляции, что чем больше липидных гранул содержится в ооцитах и эмбрионах, тем хуже эти эмбрионы и ооциты переносят процедуру криоконсервации. Однако механизмы, приводящие к гибели клеток, в результате фазовых переходов при криоконсервации и их связь с особенностями липидного состава эмбрионов и ооцитов в настоящее время не известны. Определение этих механизмов и разработка на базе этих знаний эффективных протоколов криоконсервации ооцитов и эмбрионов проблемных для криоконсервации видов млекопитающих является актуальной задачей. В 2017 году была исследована возможность детектирования фазового перехода липидов в замораживаемых эмбрионах мыши. Работа велась в сотрудничестве с Сектором криоконсервации (рук. С. Я. Амстиславский) ИЦиГ СО РАН. Эмбрионы подготавливались в ИЦиГ СО РАН, переносились в ячейку для измерения спектров КРС под микроскопным объективом, добавлялась необходимая концентрация криопротектора (глицерин) и далее эмбрион охлаждался по протоколу «программного замораживания» до температуры, при которой проводилось измерение КРС. На Рис. 1.3 показаны фотографии эмбриона при замораживании и последующем нагреве. Видно, что используемый протокол позволяет сохранить эмбрион в целостности.
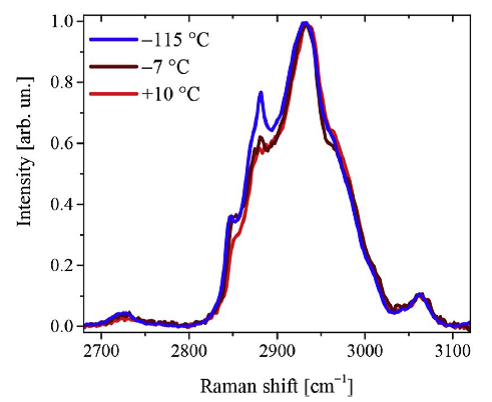
Рис. 3 Измеренные спектры КРС были скорректированы на вклад глицерина и на вклад воды в области ОН-мод. Спектры эмбриона при трех характерных температурах в области колебаний СН-мод показаны на Рис. 1.4
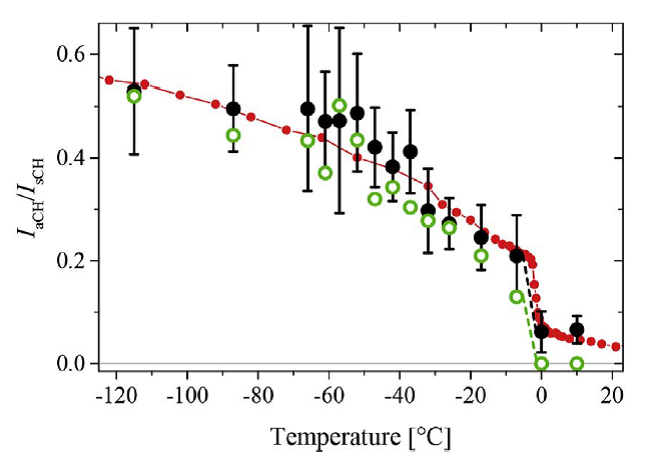
Рис. 4 Кроме вклада от СН-мод белков на рисунке 1.4 можно выделить вклад от симметричных колебаний СН2 групп липидов (2850 см-1) и от антисимметричных колебаний СН2 групп липидов (2880 см-1). Известно, что интенсивность линии КРС антисимметричных колебаний СН2 растет с ростом упорядоченности липида. Поэтому удобно рассмотреть отношение антисимметричной и симметричной линий в спектре КРС липида как меру порядка (например, Surovtsev N.V., Dmitriev A.A. // J. Phys. Chem. 2015). Отношение антисимметричной и симметричной линий в спектре КРС эмбрионов показано на Рис. 1.5 как функция температуры. Зеленые и черные кружки показывают результаты двух подходов к обработке экспериментальных данных. Красные кружки показывают как ведет себя отношение антисимметричной и симметричной линий в спектре КРС фосфолипида РОРС, где увеличение отношения с понижением температуры связано с увеличением упорядоченности конформационных состояний ацильных цепей. Прыжок параметра вблизи -2оС отражает фазовый переход гель-флюид в РОРС. Аналогично можно сделать вывод, что между 0 и +7о С в случае эмбриона мыши происходит фазовый переход липидов от разупорядоченного конформационного состояния к упорядоченному.
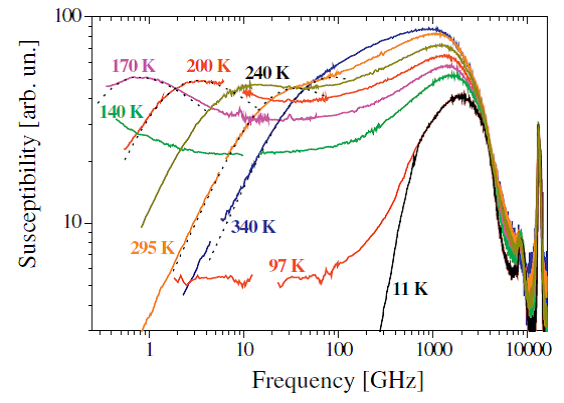
Рис. 5 3. Исследование спектров КРС и рассеяния Мандельштама-Бриллюэна (РМБ) криопротекторных растворов и стеклующихся жидкостейРанее на примере стеклующихся вандерваальсовых жидкостей был обнаружен ряд универсальных свойств материалов вблизи температуры ТА – температуры, при которой термоактивационное поведение времени релаксации (вязкости) меняется на более резкое супераррениусовское поведение. В наших экспериментах было показано, что при этой температуре зарождаются локальные короткоживущие неоднородности, которые проявляются в поведении локальной релаксации, в спектрах КРС и в поведении отношения Ландау-Плачека. Возникает вопрос – являются ли эти свойства исключительной чертой вандерваальсовских стеклующихся жидкостей или жидкости с направленными молекулярными связями также будут демонстрировать аналогичные свойства вблизи ТА? В 2017 году были исследованы спектры низкочастотного неупругого рассеяния света двух стеклующихся жидкостей глицерина и этанола, межмолекулярное взаимодействие в которых характеризуется направленной водородной связью. Для этих жидкостей была измерена зависимость отношения Ландау-Плачека от температуры и определена температура, при которой отношение Ландау-Плачека начинает отличаться от приближения однородной среды. Эта температура ассоциируется с появлением локальных неоднородностей и составила 366 К в случае глицерина и 219 К в случае этанола. Спектры неупругого рассеяния света были получены в спектральном диапазоне от 0.5 до 6000 ГГц с использованием аппаратуры для спектроскопии Мандельштама-Бриллюэна (шестипроходной тандем эталонов Фабри-Перо конструкции Сандеркока) и для спектроскопии КРС (трехрешеточный спектрометр TriVista777). Пример спектра рассеяния в случае этанола показан на Рис. 1.6 для некоторых температур в представлении восприимчивости. На температурах от 170 К и выше виден пик альфа-релаксации, который двигается в сторону высоких частот при повышении температуры.
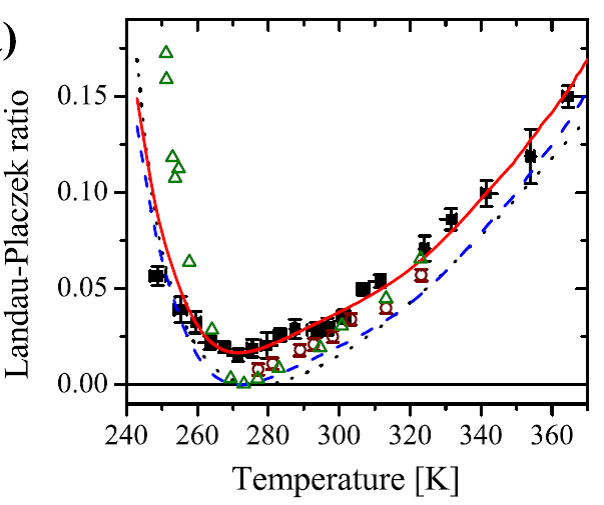
Рис. 6 Проведенный анализ спектров рассеяния света позволил определить время альфа-релаксации в широком температурном диапазоне, а деривативный анализ показал наличие перехода ТА со значениями 366 К в случае глицерина и около 200 К в случае этанола. Таким образом, стеклующиеся жидкости с водородными связями также обладают феноменом ТА – резким переходом от термоактивационного поведения к супераррениусу, сопровождающимся резким возрастанием отношения Ландау-Плачека. 4. О проблеме мгновенной структуры водыРазработанная методика определения короткоживущих локальных кластеров по отношению Ландау-Плачека была применена к проблеме мгновенной структуры воды и нами было показано, что отношение Ландау-Плачека в воде может быть описано в рамках однородной жидкости во всем исследованном диапазоне температур от -17о С и до + 90о С. Температурная зависимость отношения Ландау-Плачека в воде, полученная нами, показана на Рис. 1.7 квадратами, а красной линией показано предсказание теории однородной жидкости.
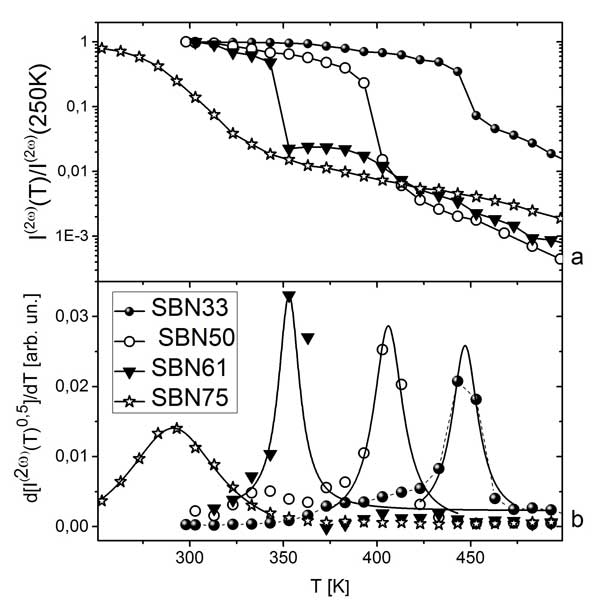
Рис. 7 5. Исследование серии кристаллов SrxBa1-xNb2O6 методами комбинационного рассеяния света, генерации второй гармоники, рассеяния Мандельштама-Бриллюэна, дифференциальной сканирующей калориметрии и диэлектрической спектроскопииВ 2017 году исследовалась серия кристаллов SrxBa1-xNb2O6 с целью прояснить особенности поведения материала вблизи сегнетоэлектрического фазового перехода. Так как, варьируя значение x в составе SBN, можно перейти от случая классического сегнетоэлектрика к сегнетоэлектрическому релаксору, это исследование может дать дополнительную информацию для актуальной проблемы описания физики сегнетоэлектрических релаксоров. Эксперименты проводились на серии кристаллов твердых растворов номинально чистых ниобата бария-стронция (SBN-x) с общей химической формулой SrxBa1-xNb2O6 (x=0.33, 0.50, 0.61, 0.75), которые были выращены способом Степанова в Институте общей физики РАН. Для исследования температурной зависимости генерации второй гармоники (ГВГ) в температурном диапазоне 200−900 K образец помещался в азотный криостат Linkam, температура в котором поддерживалась с точностью 1 K. В качестве источника облучения использовался импульсный лазер с длиной волны 1064 нм, частотой следования импульсов 1 кГц, длительностью импульса 0.6 нс и средней мощностью 100 мВт. Рассеянный назад сигнал второй гармоники через коллиматор попадал на щель спектрометра TriVista777 со спектральным разрешением не хуже 1 см-1. Спектры комбинационного рассеяния света регистрировались на спектрометре TriVista777. Для определения интегральной интенсивности и ширины сигнала ГВГ cпектральная форма сигнала подгонялась функцией Лоренца. Определенная таким образом ширина спектрального отклика на полувысоте не зависит от температуры и не превышает 1 см-1. Интегральная интенсивность данного отклика значительно превышает сигналы, которые могли бы наблюдаться в данном спектральном диапазоне и иметь другую спектральную форму. Доминирование относительно узкого и неизменного по ширине (Υ0 < 1 см-1) пика ГВГ, на фоне которого гиперрэлеевское рассеяние обнаружить не удалось, свидетельствует о том, что генерация сигнала на удвоенной частоте происходит в областях, время жизни которых превышает 5 пс. Исследование методом комбинационного рассеяния света (КРС) показало наличие центрального пика в параэлектрической фазе кристалла SBN61 с шириной, зависящей от температуры и превышающей десятки обратных сантиметров. Это свидетельствует о наличии флуктуаций параметра порядка (поляризации) в субпикосекундном диапазоне. Такие быстрые флуктуации параметра порядка происходят, по-видимому, в более крупных по размеру локальных областях, которые за времена, превышающие 5 пс, остаются асимметричными. Температурная зависимость интегральной интенсивности сигнала ГВГ, нормированная на максимальный сигнал в сегнетоэлектрической фазе, приведена на Рис. 1.8а. На рисунке виден значительный сигнал в центросимметричной фазе для всех исследованных образцов. Относительно плавное уменьшение интенсивности сигнала при увеличении температуры происходит в релаксоре SBN 75.
Рис. 8: Температурные зависимости интегральной величины сигнала второй гармоники, нормированной на максимальную величину сигнала ГВГ в сегнетоэлектрической фазе, (а) и ее производной по температуре (b) для кристаллов SrxBa1-xNb2O6 (x = 0.33, 0.50, 0.61, 0.75).
|
|
Федеральное государственное бюджетное учреждение науки Институт автоматики и электрометрии Сибирского отделения Российской академии наук |
-
Главная

-
Страницы лабораторий
-
Лаборатория 04
-
Научные и прикладные результаты лаборатории 04
- Лаб. 04 Результаты 2017